杭州师范大学生命与环境科学学院近期在《Biology and Fertility of Soils》上发表了青藏高原东部冰川消退后稀有和丰富细菌亚群的独特共生模式和驱动力的文章。在这项研究中天昊生物有幸承担了样品的相对定量扩增子测序工作。在恭喜客户又发表文章同时,我们想跟大家分享一下文章的研究思路。
英文题目:Distinct co-occurrence patterns and driving forces of rare and abundant bacterial subcommunities following a glacial retreat in the eastern Tibetan Plateau
中文题目:青藏高原东部冰川消退后稀有和丰富细菌亚群的独特共生模式和驱动力
期刊名:Biology and Fertility of Soils
影响因子: 4.829
发表时间: 2019年
研究概要:
为了深入了解丰富和稀有细菌的生态和进化过程,必须揭示它们在冰川消退中的动态和驱动力。在此,我们利用Illumina序列数据集研究了海螺沟冰川时间序列中土壤发育不同阶段相关的丰富和稀有细菌的生态丰度、演替动力学和共生模式。丰富细菌类群表现为普遍分布和紧密聚集,而稀有细菌类群则表现为沿演替阶段分布不均和松散聚集。丰富细菌和稀有细菌亚群落在装配过程中都受到不同因素的驱动:生物和土壤因子的相互作用是主要的驱动力,尽管对稀有细菌类群的重要性不如对丰富细菌类群的重要性。特别是,冗余分析和结构方程模型表明,土壤有机碳、ph和植物丰富度主要影响丰富细菌亚群,而土壤氮和pH对稀有细菌亚群落影响最大。更重要的是,与生物因子相比,土壤因子对丰富细菌(7.8%)和稀有细菌(4.5%)亚群落的影响稍大。与其它时间序列阶段相比,丰富的和稀有的细菌在中间阶段表现出更为紧凑的网络拓扑结构。在稀有类群中,重叠节点是丰富类群(Proteobacteria和Acidobacteria),和稀有类群(Planctomycetia,Sphingobacteriia, Phycisphaerae)。此外,网络分析表明,与稀有类群相比,丰富类群表现出更为密切的关系,对群落中其它共现现象的影响也更大。这些发现共同揭示了青藏高原东部冰川时间序列沿线丰富和稀有细菌亚群的不同共生模式和驱动力。
研究背景:
由全球变暖引起的冰川消退对生态系统有着深远和多重的影响,包括地表和地下的群落组成。土壤微生物是地下生态系统的独特组成部分,是土壤生物地球化学过程和生态功能的关键参与者,因此,其空间组成和分布格局的变化对环境波动的响应可能直接影响生态系统的功能。随着分子工具的发展,对微生物跨时空的生态和进化过程的研究越来越受到重视,特别是在一些严酷而原始的生态系统中,包括苔原和冰川消退区。例如,zumsteg等人认为丰富的细菌类群受冰川坝土壤水分、酸碱度和离子浓度的影响。Knelman等人发现,许多微生物在最近除冰的土壤中具有活性(或潜在的活性),能够快速利用可用的营养物质或气相沉积的富碳底物(例如花粉)。这些早期的研究主要集中在高度包容性类群(域和界水平)和环境因素对土壤群落组成的影响,而忽略了哪些较少和较不包容性类群以及微生物物种之间的相互作用。这些群落被称为稀有生物圈,丰度较低,多样性极高。Yan等人建议,为了理解微生物的演替动力学,仅分析域和界水平类群是不够的,应该同时调查包容(域和界水平)和不太包容的类群(如稀有类群),以便更好地分析群落动力学。最近基于网络分析的研究已经证明了丰富和罕见的微生物之间的相互作用在确定群落组成中的重要作用。然而,对于发展中的土壤(<200年)中微生物群落的相互作用和组装机制的了解仍然有限,特别是在冰川消退之后。
微生物群落由少数高度丰富的物种和大量的稀有物种组成,后一亚群仍有很大程度的未被开发,在大多数生态系统中,丰富的微生物有助于细菌生物量的产生和有机质的流动,而以少数个体为代表的众多珍稀物种提供了巨大的遗传多样性和功能多样性储备,可以增强丰富微生物的功能,调节生态系统的稳定性和功能。丰富和罕见的群落组合可能受到不同的控制因素的影响。然而,由于以往方法的局限性,很难直接识别稀有和丰富的微生物,也很难探索这些亚群的微生物多样性,此外,为了应对环境变化,探索稀有微生物群落的动态变化尤其具有挑战性;它们可以以非常小的规模上占据多个生态位。目前,高通量测序技术的快速发展大大提高了我们识别不同类型微生物的能力,并探索了它们在不同生态系统中的多样性和动态性。
尽管在不同的生态系统中,微生物的分布已被广泛研究,但随着时间的推移,关于微生物群落分布过程的驱动因素仍然存在争论。近年来,一些研究比较了不同环境因子对不同生态系统中丰富和稀有类群的相对影响。例如,Yan等人揭示了稀有细菌对变化条件的敏感性低于冰川消退初期丰富细菌对变化条件的敏感性。Jiao等人发现,在石油污染土壤中,稀有和丰富细菌的聚集受不同因素的驱动。稀有类群主要受随机过程的影响,而丰富的类群主要受土壤因子的控制。相反,Gobet等人发现在沿海沙地养分胁迫下,稀有和丰富的细菌类群都受到类似环境因素的影响。因此,当暴露于环境变化,特别是在发展中的生态系统中时,与丰富的类群相比,稀有类群是否表现出截然不同的生物多样性模式或不同的生态响应尚不清楚。
尽管我们知道微生物群落的多样性和组成是由环境条件驱动的,但我们对微生物类群之间复杂相互作用的理解仍然有限。一种解决方案可能是使用基于网络分析的方法,这已被证明是检验微生物群落内复杂相互作用(例如共生、寄生、竞争和捕食)的一种强有力的方法。网络的不同拓扑特征(如度、中间中心性、复杂性、模块性和聚类系数)可用于识别关键物种和评估微生物群落的稳定性。例如,高中间中心性表示网络中节点的核心和中心位置,而低中间中心性表示外围位置。此外,某些交互链接的存在或不存在可能是交互系统特定紧密性的有力指标。虽然已经开展了一些研究来研究受非生物和/或生物因素影响的微生物网络相互作用,但很少有研究将稀有和丰富类群的亚群落的共生网络模式与非生物和/或生物条件的变化联系起来,尤其是在冰川。通过下一代测序,大多数的研究集中在冰川前沿时间序列的年轻年轮序列(<200年)中的丰富细菌群落,然而,很少研究罕见生物圈和微生物群相互作用的重要性。在这项研究中,我们对脱冰层土壤中的稀有和丰富的亚群落进行了网络分析,以加深对微生物相互作用的理解。
海螺沟冰川时间序列形成的生境为研究微生物演替的多样性和轨迹提供了一个理想的自然场所,因为连续的演替阶段允许通过替代时间的空间进行详细检查。我们以前的研究表明,细菌群落受确定性过程的影响,而真菌群落则受沿冰川前场时间序列的随机过程介导。在本研究中,我们的目的是利用现有的高通量测序数据集,深入研究丰富和稀有的微生物群落,以及沿冰川前场时间序列与生物和非生物因素相关的演替轨迹和群落聚集机制。具体地说,我们假设:(1)丰富的和稀有的类群之间存在着明显的演替和组合模式,它们对群落变迁的相对贡献随时间序列的土壤发展而变得更加不均匀;(2)与稀有类群相比,丰富的类群关系更密切,对群落中其他共生物种的影响也更大,由于树种和土壤养分含量的增加,它们之间的相互作用在演替中期阶段得到加强;(3)在冰川消退的演替阶段,稀有和丰富类群装配的驱动力是不同的。综上所述,这项研究使我们能够独特地描述丰富和稀有类群的亚群落在一级演替期间如何在局部范围内发生变化,研究结果对未来全球变化下生态系统发展中的生态机制和微生物相互作用提供了新的认识。
研究方法:
采样对象
海螺沟冰川是位于青藏高原东南缘贡嘎山的季风性温带冰川。2015年8月,在海螺沟冰川年代序列中,从7个地点(代表冰消期后的不同时间:3、12、30、40、52、80和120岁)采集了土壤样本,这些地点经历了从裸露土壤到开荒群落,最终达到植被群落的最高峰。根据树木年轮和土壤侵蚀速率,采用137 CS法对各阶段的大致年龄进行了校正。在每个试验节点,建立了3个5×5平方米的试验区,试验区之间的距离为10米。用一个直径为5厘米的土壤取样器从每块地中心和4个角落采集5个20 cm深的土芯,五份土壤混成一份样本,通过2mm筛除去根。
微生物分析
16S rDNA扩增子测序(V4-V5)
研究结果:
丰富和稀有细菌类群的微生物动态变化及系统发育多样性
经过质量过滤和去除嵌合体后,扩增子测序产生1,282,898个高质量序列(每个样本44455–80714个),这些序列聚集成5584个OTU(每个样本2432±380个)。在整个数据集中,稀有类群包含1043个OTU,每个样本平均占总OTU的25.1%,但只占有0.4%的序列。只有6.7%的OTU属于丰富的微生物群,而它们占每个样本中的大部分序列(平均63.7%)。与丰富的类群相比,稀有的类群显示出更高的α-多样性(例如,OTU丰富度和香农指数)。
将一个样本中稀有或丰富的类群的占比转化成丰富、中等、稀有或未检测出,从而可视化了土壤样本中稀有和丰富类群的分布(图1)。在局部丰富的OTU中(图1a、b),32.25%的OTU在样本间也很丰富,而10.50%的OTU在其它样本中很稀有。在局部稀有的OTU中(图1c,d),52.38%的OTUs在其它地方未检测到,19.26%为中等丰度,几乎没有OTUs在其它样本中是丰富的。
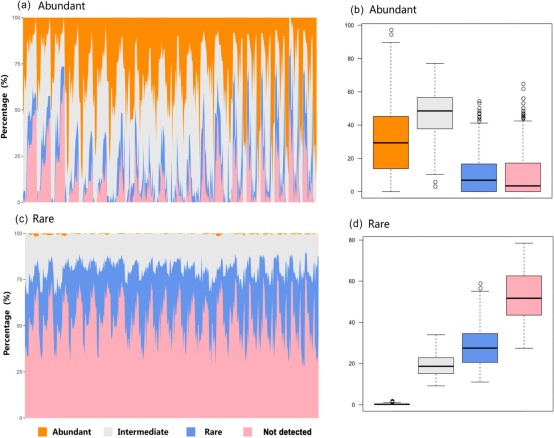
图1海螺沟冰川时间序列中丰富和稀有菌的分布。面积图显示了一个样本中丰富(A,B)或稀有类群(C,D)的百分比变化。
丰富和罕见的亚群落成分之间存在差异:Alphaproteobacteria, Betaproteobacteria, Sphingobacteria, Acidobacteria_Gp6, Acidobacteria_Gp3, Acidobacteria_Gp2, Acidobacteria_Gp1, Acidobacteria_Gp4是最丰富的类群,Gammaproteobacteria, Deltaproteobacteria, Planctomycetia, Cytophagia, Chlamydia是最稀有的类群。微生物群落在时间序列上具有系统学上的不同(图2)。整个群落和丰富细菌的NRI(净种间亲缘关系指数)值随着土壤年龄的增长而增加,而NTI(净最近种间亲缘关系指数)值则下降(图2a-e)。然而,对于稀有细菌来说,随着土壤年龄的增长,NTI和NRI都显著降低(图2c,f)。
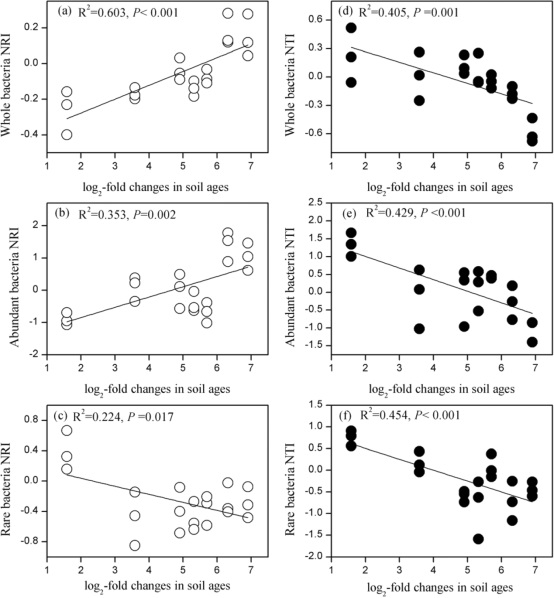
图2 海螺沟冰川时间序列各阶段(年龄)整体群落,丰富群落和稀有群落净种间亲缘关系指数(NRI)(A-C)和净最近种间亲缘关系指数(NTI)(D-F)的回归分析
不同亚群的共现模式与拓扑性质
元群落共现网络在3728个OTU(节点)中捕获了293352个关联(边缘)(图3a)。此外,361个节点和218个节点分别属于丰富类群和稀有类群。与稀有类群(82个内部关联)相比,丰富类群(111014)和中间丰富类群(117110)的内部关联更为复杂。对于外部关联,我们观察到丰富类群和中间丰富类群节点之间的关联数量最多(63411),稀有类群和丰富类群节点之间的关联数量最少(23)。这些观察表明,中间类群经常与丰富类群共存,而稀有类群很少与丰富类群共存。在核心分类群和其他分类群的网络中(图3b),分别有8126和131664个内部关联,核心类群和其他类群之间有53562个外部联系(图3b)。此外,四个拓扑特征(度、中间性、紧密性和特征向量中心性)的值,对于丰富类群比稀有类群更高,对于核心类群也比其他类群更高。在不同的演替阶段生成微生物共现网络,计算网络拓扑特征,探讨微生物对土壤发育的响应。在时间序列的中间阶段,网络变得最为聚集和复杂,其节点、边缘关联和内部和外部关联的数量最多。
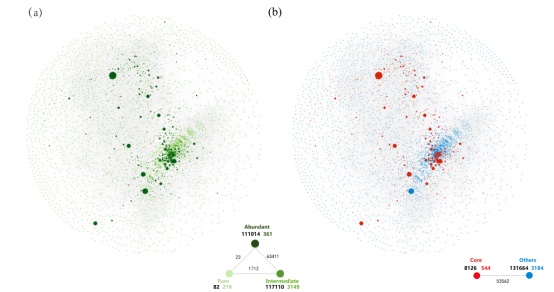
图3 基于相关分析的海螺沟冰川时间序列中丰富稀有细菌共生网络图:由丰富或稀有的分类群(A)和核心或其他分类群(B)组成的共生网络。节点大小与OTU的相对丰度成正比,节点之间连接的厚度(即边缘)与Spearman相关系数的值成正比。每个生物圈之间的内部和外部关联(黑色数字)显示在每个图的右下角,每个生物圈中节点号的颜色对应于类别。
不同微生物亚群落的模块结构
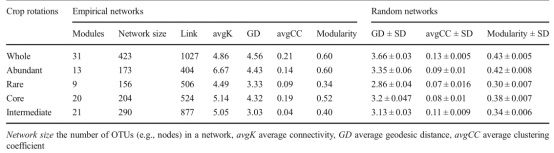
根据系统发育分子生态网络计算的模块,我们发现整个群落以及丰富亚群、稀有亚群、核心亚群和中间亚群被清晰地分为31、13、9、20和21个主要模块(表1)。与随机网络相比,经验网络具有更高的avgCC、平均GD和模块化值(表1),此外,与稀有亚群子网(分别为0.09和4.49)相比,丰富亚群子网具有更高的avgCC(0.14)和平均连通性(6.64),这意味着稀有亚群子网比丰富亚群子网更分散,连接更少(表1)。此外,模块中心和连接分布在各种微生物类群中,我们在所有细菌的网络中检测到11个模块中心,其中3个与Acidobacteria有关,1个与Alphaproteobacteria有关,3个与Bacteroidetes有关,而其它的则属于Alphaproteobacteria, Betaproteobacteria, Planctomycetia, Actinobacteria, Omnitrophica。此外,我们还检测到四个连接,分别属于Chloroflexi, Gemmatimonadetes, Proteobacteria, Actinobacteria。
表1冰川时间序列上整个群落以及丰富亚群、稀有亚群、核心亚群和中间亚群的经验分子生态网络及其相关随机分子生态网络的拓扑特征
此外,三元图分析表明,大多数与Proteobacteria, Bacteroidetes, Acidobacteria相关的OTU在所有连续阶段都很常见(图4),Actinobacteria和 Gemmatimonadetes主要在早期和中期富集,而Planctomycetes和Verrucomicrobia则在晚期富集(图4)。
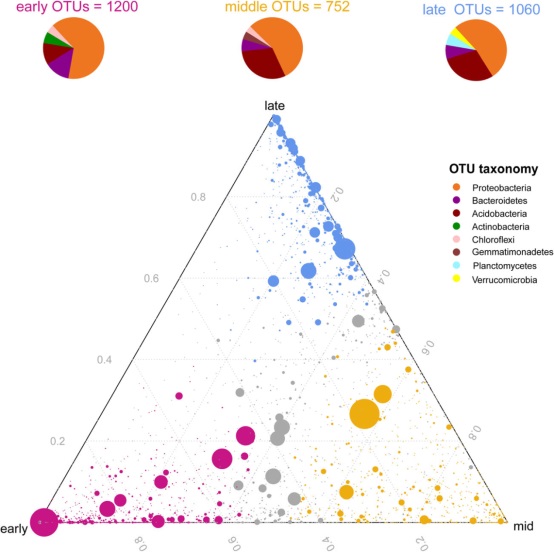
图4 三元图显示了三个连续阶段(3-120年)的早期、中期和晚期所有OTU的相对丰度(>5‰)。每个点代表一个OTU。每个圆的大小代表它的相对丰度。它的位置代表了它相对于每个隔间的相对丰度。不同颜色的圆圈表示一个隔间内显著富集的OTU(例如,晚期为蓝色,中期为橙色,早期为洋红),而灰色圆圈表示在特定隔间内没有显著富集的OTU。
丰富亚群与稀有亚群的反应与驱动力对比
五个亚群的β-多样性在时间序列阶段表现出明显的不同反应(图5a)。稀有亚群的β-多样性明显高于其它四个亚群。丰富亚群和核心亚群的β-多样性明显低于整个群落(图5a)。将微生物β-多样性划分为丰富度和更替(物种替换)组分表明,物种更替对β-多样性的贡献高于各亚群的物种丰富度(图5b)。
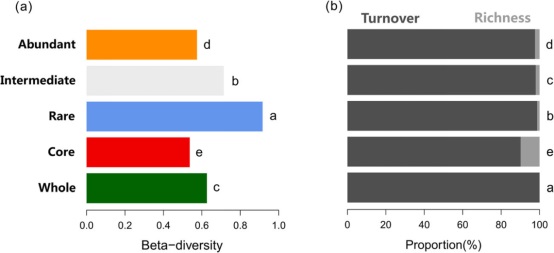
图5 基于Bray-Curtis(a),和物种更替以及丰富度的相对贡献(b)进行的海螺沟冰川年代不同细菌亚群β-多样性分析
根据NMDS图,将不同演替阶段的细菌群落(包括所有物种群落、丰富物种和稀有物种)分为三个类群:S1作为类群1,S2-S5作为类群2,S6-S7作为类群3(图6a-c)。与稀有亚群相比,丰富细菌亚群的格局更接近于整个细菌群落的格局。从不同演替阶段的松散聚类和不同演替阶段的群间距离比较可以看出,稀有类群与丰富类群及所有类群相比具有显著的分散性(图6c、f),这些观察结果得到了稀有亚群和丰富亚群RDA结果的证实,表明细菌群落按演替阶段强烈聚集。
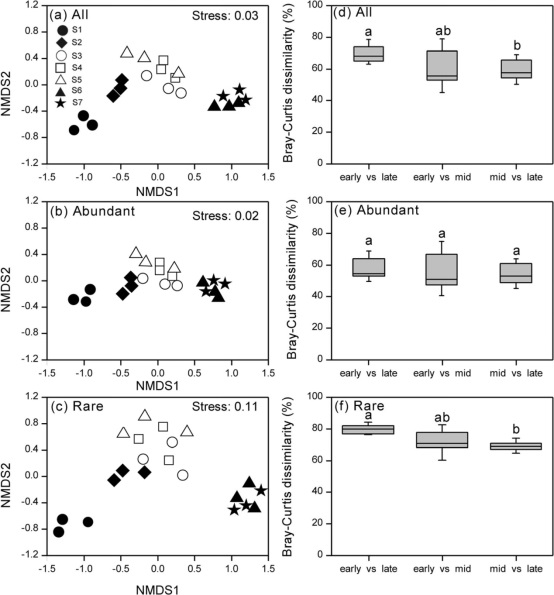
图6 海螺沟冰川时间序列细菌群落比较。 基于Bray-Curtis距离(A-C)的细菌群落非度量多维标度(NMDS)排序。不同演替阶段(d-f)土壤微生物群落的Bray-Curtis差异分析。
此外,前两个RDA组成部分解释的总变化,对于丰富亚群(73.9%)要比稀有亚群(19.43%)高得多(图7a,b)。变异分配分析表明,生物和土壤因子的相互作用比土壤和生物因子单独在确定丰富和稀有亚群组成中更为重要(图7c,d)。此外,扫描电镜模型显示,土壤年龄、土壤酸碱度、土壤有机碳和植物丰富度是丰富类群多样性的重要因素,而土壤年龄、土壤酸碱度、总氮和土壤有机碳是稀有类群多样性的重要因素(图7e、f)。
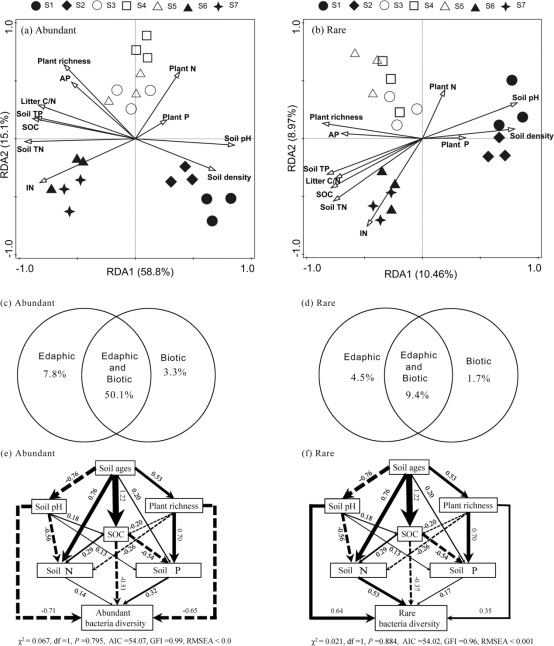
图7海螺沟冰川时间序列上丰富和稀有亚群结构的所选环境变量的冗余纵坐标(a,b)、变异分配分析(c,d)和结构方程建模(e,f)。