DNA甲基化是一种表观遗传标记,它参与了包括植物发育、胁迫适应和基因组进化等生物过程。DNA甲基化作为一种新的遗传变异来源,它在增强植物抗病能力方面具有巨大潜力,为科学家和育种者带来了新的方向。今天介绍的这篇综述就系统的讨论了病原体诱导的DNA甲基化修饰是如何影响宿主的转录重编程和基因稳定性的,以及DNA甲基化在植物病原体相互作用中的功能。这些都将有利于育种者提高作物抗性和减少产量损失。

发表期刊:Trends in Plant Science 发表时间:2019-10-8 影响因子:14.006
植物病害与表观调控
植物病害导致全球产量大幅下降。例如由Magnaporthe oryzae引起的稻瘟病正在全球范围内使水稻产量减少30%。传统的作物病害管理策略,如杀菌剂施用、作物轮作和种子健康监测不足以自行控制产量损失。减少由疾病引起的产量损失的更可行的方法是将传统策略与分子植物育种方法相结合,这种方法依赖于抗性基因(R基因)的遗传多样性驱动的表型多样性,抗性基因包括品种特异性抗性基因和参与品种非特异性抗性的基因。
最近,表观遗传的变化也被证明在诱导表型多样性方面也有重要作用,包括通过控制防御基因的表达水平来诱导植物抗性反应。表观遗传变化包括DNA甲基化和去甲基化、染色质重塑和组蛋白修饰等。在真核生物中,DNA甲基化是一种重要且常见的表观遗传修饰,涉及多种生物过程,包括植物对胁迫条件的适应机制、基因组管理和发育过程。
植物防御机制与DNA甲基化
植物利用不同层次的防御机制来抵御病原体。相比之下,病原体使用不同的生活方式和策略与宿主植物相互作用。病原体感染策略的例子包括细菌在细胞间区的增殖,线虫直接从植物细胞中取食,真菌菌丝进入植物细胞或其周围的扩张,以及病毒在植物细胞中的存活和增殖。植物对抗病原体的能力依赖于防御基因的调控,包括植物防御机制第一层中的基因,这些基因通过表面宿主蛋白受体识别病原体/微生物效应子、病原体相关分子模式(PAMPs)或微生物相关分子模式(MAMPs)而启动。这些基因刺激第二层防御机制中的PTI或ETI机制。ETI是一个抗性基因,其中抗病蛋白和特异性病原体毒力(Avr)蛋白之间的相互作用。NLR蛋白是最著名的抗性蛋白,对植物抗病性有重要影响。就数量和分布而言,这些基因的表达模式和进化是植物胁迫诱导反应的关键因素。
DNA甲基化是指在DNA的胞嘧啶碱基上增加一个甲基,形成5-甲基胞嘧啶。在植物中,胞嘧啶碱基的甲基化是在对称CG和CHG以及不对称CHH(其中H = A、C或 T)上。甲基化主要发生在CG上,然后是在CHG和CHH,包括拟南芥(CG: 24%, CHG: 6.7%,CHH: 1.7%),木薯(CG: 58.7%, CHG: 39.5%, CHH: 3.5%),大豆(CG: 63%, CHG: 44%, CHH: 5.9%),玉米(CG: 65%, CHG: 50%, CHH: 5%)和水稻(CG:54.7%, CHG: 37.3%, CHH: 12%)等。上述研究还表明,基因体(Gene body)甲基化在CG上发生率很高;而转座因子(TEs)(即一种类型的移动遗传因子)在所有三种序列中都高度甲基化。
植物中的DNA甲基化涉及三个过程: DNA甲基化从头生成、甲基化的维持和DNA去甲基化。在植物中, DNA甲基化从头生成是由DNA重排甲基转移酶2(DRM2),一种哺乳动物DNA甲基转移酶(Dnmt3)的同系物催化的。从头生成甲基化通过小RNAs发生,称为核RNA介导的DNA甲基化(RdDM)机制,需要两种植物特异性RNA聚合酶,Pol IV和 Pol V。小RNAs包括siRNA和miRNA,已被证明在包括基因表达调控和转座子沉默等一系列生物过程中具有重要作用。例如,已经证明miRNAs参与调控番茄中NLRs的表达,进而调控抗性反应。还发现多种水稻miRNAs通过上调防御相关基因参与增强对水稻稻瘟病的抗性。在拟南芥中,作为siRNA介导的沉默中导致DNA和组蛋白甲基化的主要成分之一的Argonaute 4 (ago4)的突变,导致对丁香假单胞杆菌的易感性的增加。拟南芥RdDM途径多个突变体抗病性的改变,包括nrpe 1(Pol V的最大亚基)、nrpd 2(Pol V的第二大亚基)、ago4、drd1、rdr2(依赖RNA的RNA聚合酶2),都证实了RdDM对植物免疫系统是至关重要的。
在植物中,最丰富的一类小RNAs来源于TEs和DNA重复序列,含有TEs的基因更容易通过RdDM途径被调控。非TE相关基因的表达也可能受到RdDM途径的影响。为了平衡基因组的甲基化水平并维持基因表达,植物使用DNA去甲基化酶去除5-甲基胞嘧啶。在拟南芥中有四种DNA去甲基化酶:DME、ROS1、DML2和DML3 (图1)。
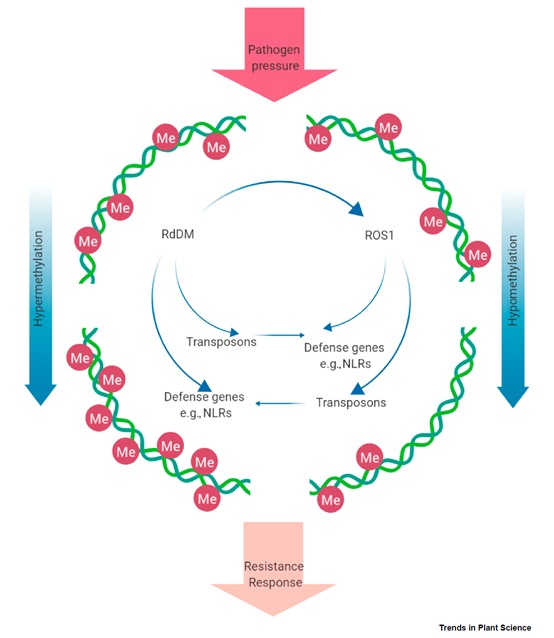
图1、病原菌胁迫下植物的DNA甲基化调控。病原菌攻击期间植物通过RNA介导的DNA甲基化(RNA-directed DNA methylation, RdDM)造成的甲基化程度升高和沉默1阻遏物(Repressor of Silencing 1, ROS1)导致的甲基化程度下降可以调控如NLR等防御基因的表达,并修饰转座子活性。转座子活性的变化也会影响防御基因的表达。ROS1的表达也可以通过RdDM途径调控。
DNA甲基化的维持取决于类型。CG甲基化的维持是通过Methyltransferase1(MET1)进行的,在CHG中,甲基化的维持是通过Chromomethylase3(CMT3)和CMT2进行的,CHH甲基化的维持是通过CMT2或RdDM进行的。拟南芥缺失甲基化维持的突变体,显示对丁香假单胞杆菌的抗性增强。
随着检测基因组DNA甲基化的多种方法的迅速发展,也为植物生物发育等过程中DNA甲基化的研究,包括植物病原体相互作用中的作用提供了技术支持。这些方法包括whole-genome bisulfite测序、epi-restriction-site associated DNA测序、methylC-capture测序,methylated DNA immunoprecipitation测序、combined bisulfite restriction分析、SeqCap Epi,以及amplification of intermethylated sites等方法。表1总结了应用这些不同方法对作为病原体诱导的植物免疫系统DNA甲基化进行的最新研究。显然,在植物中,在病原体胁迫下,DNA甲基化是高度动态的,可以显著影响抗性反应(图1)。
表1、病原体胁迫下DNA甲基化修饰对植物的影响

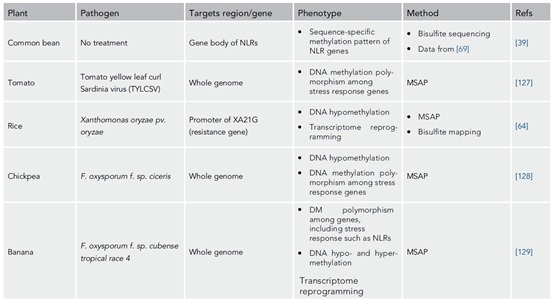
注:AFLP, amplified fragment length polymorphisms; COBRA, combined bisulfite restriction analysis; HPLC, high-performance liquid chromatography; LRR, leucine rich repeat; MSAP, methylation-sensitive amplified polymorphism; MeDIP-seq, methylated DNA immunoprecipitation sequencing.
DNA甲基化:对转录组重编程的刺激
植物对逆境的适应依赖于转录组的重编程,并且在不同的生长阶段和逆境条件下,DNA甲基化在转录组的改变中起着至关重要的作用。DNA甲基化和基因表达之间的关系非常复杂,取决于各种因素,包括组织类型、转座子活性、序列背景和基因组区域,例如基因体和启动子。
1、启动子甲基化
许多研究表明启动子区域的甲基化调控基因的表达,甲基化启动子和基因表达水平之间呈负相关。致病相关基因启动子(如NLR基因)中病原体诱导的DNA低甲基化可以改变它们的表达水平并诱导抗性反应。例如,水稻白叶枯病菌抗性基因Xa21G的启动子区,在水稻突变体中低甲基化,在野生型植物中高甲基化。随后,在突变植物中观察到高水平的基因表达和对病原体的抗性表型,但在野生型中没有观察到。与生物胁迫一样,由非生物胁迫(如盐和干旱)导致的启动子低甲基化,使得非生物胁迫应答基因表达上调。然而,启动子的低甲基化并不总是基因表达水平增高所必需的。例如,尽管抗稻瘟病基因Pib在感染稻瘟病菌的植物中有高水平的表达,但是它的启动子是高度甲基化的;有趣的是,Pib启动子的部分去甲基化反而降低了它的表达。
在胁迫条件下,低甲基化和高甲基化对植物来说可能都是有益的,因此高水平的全基因组甲基化会降低转录,从而降低细胞的能量消耗,在病原体攻击或非生物胁迫期间降低能量消耗是必要的。相比之下,抗性基因的低甲基化会增强它们的表达,从而对环境因素产生快速的适应性反应。为了最大限度地利用DNA甲基化来提高抗病能力,了解植物对病原体采用的策略是低甲基化还是高甲基化是很重要的。
2、基因体甲基化
与启动子甲基化在基因表达中的作用相反,基因体甲基化在基因表达修饰中的作用仍不太清楚。通常,在植物和哺乳动物中,转录/编码区甲基化水平高的基因常为组成型高水平的表达。对拟南芥全基因组的DNA甲基化分析表明,中度转录的基因最有可能甲基化,而表达水平最低或最高的基因最不可能甲基化。基因体中CG甲基化已被证明与许多植物物种中的基因表达呈正相关,包括木薯、大豆、水稻和拟南芥。相比之下,在番茄、拟南芥、豆子基因体中的CHG和CHH甲基化与基因表达呈负相关。这些观察表明基因体甲基化的序列背景对基因表达调控很重要。
除了序列背景,有证据表明基因体甲基化对基因表达的影响取决于甲基化位点的位置(即内含子、外显子和它们的边界)。在拟南芥中,CHH和CHG甲基化在很大程度上被排除在基因体之外,内含子-外显子边界区除外。内含子、外显子和它们的边界中不同的DNA甲基化模式可以控制RNA剪接事件,通过产生新的功能性或功能失调性基因转录物,甚至沉默基因,导致表型多样性增加。
3、TE甲基化
TEs通过依赖于TEs激活的各种机制调控基因表达,而TEs激活主要由它们的DNA甲基化状态控制。全基因组的低甲基化是生物或非生物胁迫的结果,可以激活转座子,增加它们在病害相关基因中的流动性,并随后调控它们的表达水平。
TEs用来调控基因表达的这些机制包括:(I)插入基因区域,改变表达水平或沉默基因;(ii)启动子的插入/缺失和新启动子的产生;(iii)启动子中已经存在的TEs的DNA甲基化状态的改变;和(iv)基因的上游或下游插入,改变它们的表达。虽然这四种机制的潜在过程在植物病原体的相互作用中还没有被很好地解释,但是有一些例子可以证实宿主TE甲基化模式受到病原体的影响,并且在调控防御机制中起重要作用(表1)。另外,抗性基因启动子中TEs的甲基化模式已被证明控制抗性表型和产量损失。TEs的甲基化模式可能是同时调控各种基因的关键因素,因此,管理TEs的甲基化模式可能是育种计划中提高理想农艺性状表达和减少不理想农艺性状影响的一种途径。
在水稻中,编码转录因子的WRKY45的两个等位基因在抗水稻白叶枯病中具有相反的功能,强调了基因甲基化在基因功能调控中的重要性。两个等位基因之间的区别是WRKY45-2等位基因中存在两个MITEs,而WRKY45-1等位基因中不存在它们。这些MITEs参与小RNA(TE-siR815)的产生,TE-siR815通过RdDM途径提高一种NLR基因ST1内含子的DNA甲基化水平,从而抑制ST1的表达,解释WRKY45-1在调控抗病性中的负作用和WRKY45-2的正作用。
DNA甲基化:基因组稳定的力量
基因组稳定性取决于TEs的激活和基因组自由的重排;活跃的移动元件和高比率的基因组重排导致基因组不稳定。DNA甲基化对基因组稳定性的影响不如转录组重新编程的作用那么快,并且长期的由于各种环境刺激导致的DNA甲基化模式的改变,也会影响基因组进化过程。
抗性基因变异的主要进化机制是通过全基因组复制和三倍化、转座子介导的基因复制或串联和片段重组。DNA甲基化,以及其他表观遗传标记,在控制这些基因组进化过程中起着重要作用。例如,在大豆中,复制基因中基因体CG甲基化很丰富,与未甲基化基因相比,其进化速度也较慢。同样,在水稻和木薯中已经显示了DNA甲基化对重复基因进化的影响。有趣的是,在小麦中Ppd-B1的拷贝数变异(CNV)与其DNA甲基化水平之间存在正相关,因此高甲基化株系比低甲基化株系拷贝数多。这一观察表明,在Ppd-B1位点高水平的DNA甲基化可能是CNV形成的一个调控因素。
由于暴露于胁迫下基因组DNA甲基化状态的突然变化会加速这些进化过程的发生,由此DNA低甲基化促进暴露于病原体的组织中体细胞重组的增加。病原体攻击对宿主基因组稳定性的影响,主要受TE活性、重排过程和迁移的影响 (图2)。
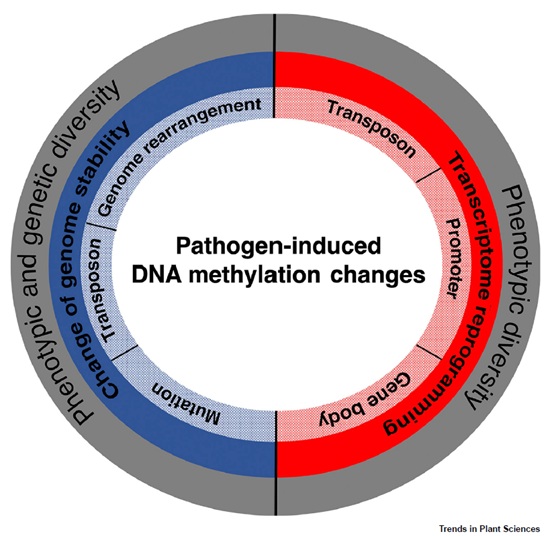
图2、病原体诱导的DNA甲基化变化及对通过转录组重编程和基因组稳定性变化对表型和遗传多样性的影响。
1、TE甲基化
基因组使用DNA甲基化和其他表观遗传标记来沉默TEs并限制它们的流动性,以增加基因组稳定性;然而,在胁迫下TEs的激活有利于植物的适应。TE插入编码区可以产生突变,增加遗传多样性,从而增加表型可塑性,这是植物在动态环境中所必需的。例如,在拟南芥中,由热应激激活的ONSEN及其插入ABSCISIC ACID-INSENSITIVE基因产生突变,导致脱落酸不敏感表型,进而导致胁迫耐受性。
基因组也利用DNA甲基化来控制DNA插入位点,并最大限度地减少DNA移动的负面影响。在玉米中,在高表达基因的起始、终止和内含子区观察到较低的CG甲基化率;相比之下,外显子在CG中高度甲基化。在基因体中具有优先插入位点的转座子Mutator(Mu)的甲基化模式显示,其插入位点区缺乏CG但确富含CHG和CHH。这一观察表明,玉米基因组通过CG甲基化保护编码区,并使内含子无突变。这种位点特异性的DNA甲基化引导DNA插入基因组的不同部分,包括基因的编码区和非编码区,这可以增加选择性剪接事件的机会,创造新的基因,增加表型和基因型可塑性。这些观察表明,TE活性和基因组不稳定性对基因组没有任何危害。
2、基因组重排
胁迫条件下的表观遗传变化增加了非同源末端连接和同源重组(HR)的DNA修复,从而降低了基因组稳定性。尽管胁迫条件可能不会直接损害DNA,但它们会增加基因组重排过程的发生。例如在非最佳温度和日长增加下的拟南芥中,在病原体胁迫下的拟南芥和烟草中,在UV-B和UV-C辐射下的拟南芥中,以及在盐胁迫下的拟南芥中,其HR频率会增加。宿主植物和病原体都使用DNA甲基化来控制基因组重排过程,以增加它们自身的表型和遗传多样性。
胁迫诱导的表观遗传变化激发的基因组重排过程显示了世代间的稳定性和可遗传性。例如,在用UV-C或鞭毛蛋白处理的拟南芥植物中,表观遗传变化在至少四代中是稳定的;在用TMV感染的烟草中,两代以上表现出HR频率的增加;在用甲基化抑制剂处理的水稻种子中的表观遗传变化可以遗传九代以上。然而也有人认为,维持多代的效应依赖于反复暴露在胁迫下,这些表观遗传变化只在一代中是稳定的,因此表观遗传变化的遗传性还需要进一步研究。
3、突变
甲基化胞嘧啶残基是自发脱氨的热点。甲基化胞嘧啶和鸟嘌呤的脱氨基作用将它们分别转化为胸腺嘧啶和腺嘌呤。这些脱氨基作用是植物、人类和微生物基因组突变的主要来源之一,并导致了几种人类遗传性疾病。甲基化水平高的小麦祖先显示出比甲基化水平低的祖先更高的C:T或G:A转换速率,暗示基因组突变速率可以由基因组甲基化水平驱动,其中C:T或G:A转换是5-甲基胞嘧啶脱氨的典型结果。结果表明祖先甲基化控制了后代基因组多态性的水平。
很明显甲基化胞嘧啶向胸腺嘧啶的转换受环境因素的显著影响。从C:G到 T:A转换速度与温度的升高成正相关,在数十亿年的进化过程中,由于地球表面的冷却,这些突变的速度已经下降。哺乳动物的阳光照射也显示出转换的速率的增加。温室中拟南芥植物超过30代的突变谱显示,大多数自发突变是G:C到A:T的转换,主要是甲基化胞嘧啶脱氨和紫外线诱导诱变的组合效应。但目前没有研究支持在病原体压力下的植物中的这些情况产生。
结论及未来方向
DNA甲基化是植物适应的一种机制,是一种相对新发现的变异来源。植物利用DNA甲基化来指导长期生物过程(如基因组进化)和短期过程(如对胁迫的反应)。全基因组的低甲基化和高甲基化,响应环境因子,可以改变基因组的稳定性并诱导进化过程,如突变和片段重排,从而产生稳定的遗传和表型多样性。
在突然的胁迫暴露下,通过调控胁迫反应基因的表达水平,DNA甲基化的改变作为一种快速的适应性反应。基因上游和下游、TEs和基因体的低甲基化和高甲基化主要影响转录过程。总之,植物利用DNA甲基化修饰来扩大它们在动态环境中的表型和遗传多样性,以便在需要时最大限度地提高能量消耗效率和抗性反应。
从育种者的角度来看,植物病害管理和抗性提高一直是育种规划的重中之重。但是气候变化和人口增长带来了额外的挑战。由于气候变化,胁迫和疾病流行的严重性和频率增加,意味着产量损失预计将显著增加。加上人口增长率和未来粮食需求的增长趋势,这给育种者施加了更大的压力,以尽量减少产量损失。育种者需要超越目前主要依赖于遗传变异的育种策略,以最大限度地减少因植物病害造成的产量损失。通过扩大表型变异,DNA甲基化可以解决育种计划中存在的局限性。本文研究者认为,DNA甲基化在植物病原体相互作用中的功能值得进一步研究,以填补关键的知识空白,并使其能够应用于提高抗性的育种计划中。
昊技术:
天昊生物提供全方位DNA甲基化检测服务,尤其在目标区域DNA甲基化检测方面优势明显。天昊专利MethylTarget®多重目的区域甲基化富集测序技术,在大幅度降低研究费用的同时实现个性化目的区域测序检测。
技术优势:
• 基于二代测序,可以获得目标区域内所有C的甲基化数据。
• ~500×的测序深度,精确计算每个位点C的甲基化程度。
• 通量高,可同时对成百上千个区域进行甲基化程度分析。
• 适用于大样本多区域的DNA甲基化水平检测。
想要马上开始项目检测吗?欢迎联系我们具体咨询!邮箱:techsupport@geneskies.com
电话:400-065-6886