题目:Analysis of genetic diversity of ancient Ginkgo populations using SSR markers
发表期刊:Industrial Crops and Products 发表时间:2019-11-18 影响因子:4.191

银杏(Ginkgo biloba L.)是一种著名的孑遗植物,也是银杏纲唯一幸存的物种。为了探讨银杏地理种群的遗传变异和冰川期可能避难所,研究者利用22个SSR标记对6个古种群和2个栽培种群的216份样品进行了分析。在22个SSR位点共检测到231个等位基因,平均预期杂合度(He)为0.808,表明遗传多样性水平较高。STRUCTURE、系统发育树和主坐标(PCoA)分析结果表明,8个种群可分为2个类群,即东部类群(TM、NJ和PZ)和西南部类群(PX、DY、MJ、FG和WC)。东部组和西南部组分别有15个和14个独特的等位基因。在这些群体中,来自东部的群体TM具有最高的遗传多样性和等位基因丰富度,在该群体中检测到9个稀有等位基因和8个特异等位基因。虽然西南地区WC群体的遗传多样性低于TM群体,但在WC群体中发现了8个稀有等位基因和2个特异等位基因。据推测,东部的群体TM地区和西南部的群体WC地区是冰川期的避难所。此外,在来自东部的2个栽培种群中总共检测到7个特异等位基因,这表明在冰川期期间可能还有其他避难所。
1、研究背景
银杏作为一种“活化石”,是银杏属中唯一的物种。银杏不仅是一种非常重要的药用植物,还具有木材和观赏用途。此外,银杏因其起源早、生命力强,对植物物种起源和进化的科学研究非常有价值。然而,关于银杏的起源,没有确凿的证据或公认的结论。侏罗纪时期银杏科物种几乎遍布全球,然而,随着白垩纪晚期地球气候的剧烈变化,银杏树属物种开始减少。到第四纪冰川期开始时,冰川运动已经导致北美和欧洲所有银杏灭绝。幸运的是,由于特殊的地形特征,中国的一些地方由于冰川侵蚀,成为银杏的独特避难所。然而,这些庇护所的数量和地点尚未得到确认。
在本研究中,研究者调查了分布在两个潜在避难所的银杏资源,并收集了古代种群。结合两个人工林,利用SSR标记(1)了解这两个潜在保护区的遗传多样性和种群间的关系;(2)探索银杏在中国的可能起源和可能保护区。
2、材料和方法
植物材料
在中国西南部发现了五个古代种群(PX,DY,MJ,FG和WC),而在中国东部只发现了一个古代种群(TM)。取样树之间的距离超过50 m,以降低取样树相关的概率(小种群除外)。为了比较古老种群和栽培种群之间的遗传差异,从中国东部共选出两个栽培种群(NJ和PZ)。总共从6个古代种群和2个种植园收集了216个个体。收集幼叶,用硅胶干燥,在-80℃保存备用。每个群体的详细信息见表1和图1。
表1、银杏种群相关信息
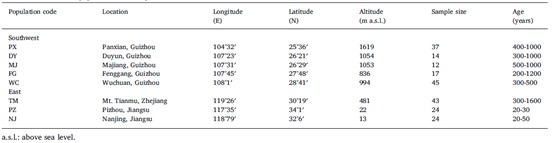
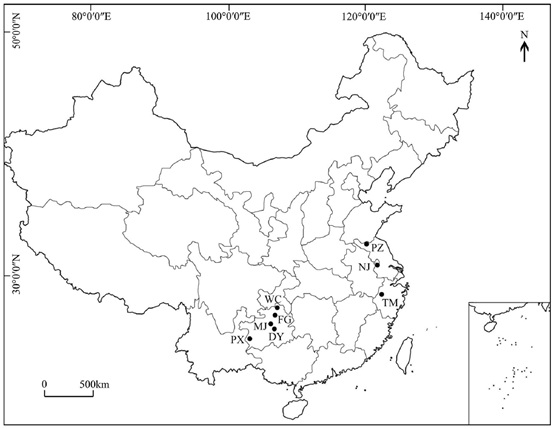
图1、银杏8个种群的地理分布。PX,DY,MJ,FG,WC和TM是古代种群,NJ和PZ是栽培种群。
DNA提取及SSR标记基因分型
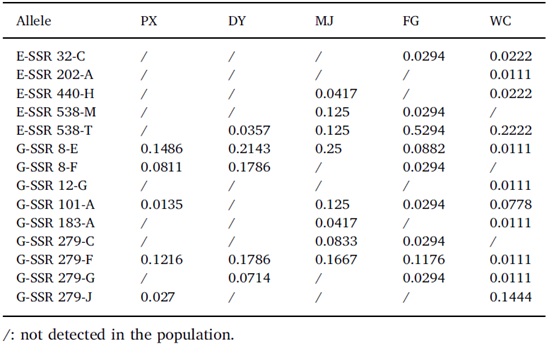
使用CTAB方法提取总基因组DNA。根据扩增的多态性、稳定性和清晰度,从本实验室开发的556个SSR标记中选择了22个SSR标记,包括8个表达序列标记(E-SSR)和14个基因组标记(G-SSR) (表2)。使用ABI Veriti 96 PCR system (Thermo Fisher Scientific, MA, USA)对所有标记进行PCR扩增。使用96个毛细管3730xl DNA分析仪分离反应产物。
数据分析
对于每个标记,使用Peak Scanner软件(Thermo Fisher Scientific, MA, USA)分析基因型结果,具有相同碱基大小的条带用相同的大写字母表示。使用POPGENE软件测定观察等位基因数(Na)、有效等位基因数(Ne)、观察杂合性(Ho)、预期杂合性(He)、Nei多样性指数(H)、遗传变异指数(Fst)、基因流(Nm)和Hardy-Weinberg平衡评估值(通过卡方检验评估)。使用PowerMarker软件进行多态性信息含量(PIC)的测定和基于neighbor-joining方法构建的系统发生树。使用FSTAT软件分析等位基因丰富度。分子方差分析(AMOVA)和主坐标分析(PCoA)使用GenAlEx软件进行。
为了估计个体祖先,本文使用STRUCTURE软件对八个样本集(DY、MJ、FG、WC、PX、TM、NJ和PZ)进行贝叶斯聚类分析。对于每个K值(潜在祖先群体的数量,范围从1到9(群体数量+ 1)),基于混合模型估计每个个体的遗传祖先;估计是用马尔可夫链蒙特卡罗(MCMC)方法获得的,进行迭代100,000次及100,000次的burn-in。根据STRUCTURE HARVESTER中的Evanno方法确定了最佳K值。结果用CLUMPP和DISTRUCT绘制。
3、研究结果
遗传多样性
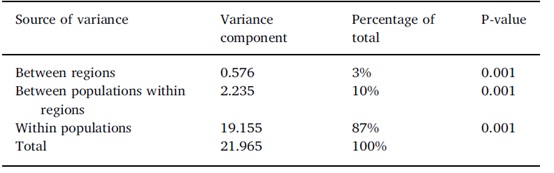
基于来自六个古老群体和两个栽培群体的216个样本,在22个SSR基因座共获得231个等位基因(图2)。每个SSR基因座的等位基因数从5到20不等,平均每个基因座有10.5个等位基因(表3)。总的来说,平均Ne、He和PIC分别为6.104、0.808和0.781,表明216个样品含有高水平的遗传多样性。在22个SSR基因座中,E-SSR32基因座遗传多样性水平最低,Ne最小(2.386),He最低(0.582),PIC最低(0.565)。相比之下,G-SSR38基因座表现出最高的多样性,因为该基因座具有最大的Ne (12.130)、He (0.920)和PIC (0.912)。然而,22个SSR位点的等位基因频率明显偏离哈迪-温伯格平衡(P < 0.05)。
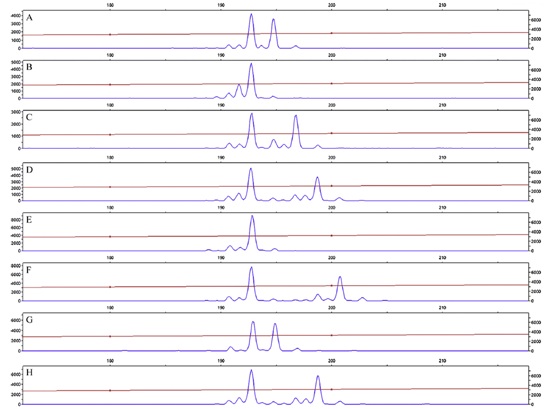
图2、E-SSR32基因座位点的PCR产物分离图谱。前四个 (从A到D)来自WC种群,后四个(从E到H)来自TM种群。
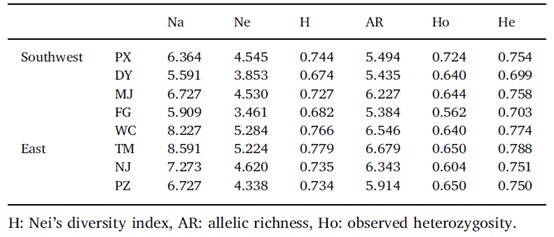
总的来说,六个古代和两个栽培群体的遗传多样性没有显示出很大不同,这些群体的遗传多样性从高到低依次为TM、WC、PX、NJ、PZ、MJ、FG和DY。在八个群体中,He值高于Ho值,表明杂合子缺乏(表4)。在6个种群中,种群TM表现出最高的遗传多样性,群体DY的遗传多样性最低。西南地区古树的遗传多样性(WC、PX、FG、DY和MJ)也是略不同于东部古树的遗传多样性(TM)。
表4、8个群体22个SSR位点遗传多样性统计平均值。
遗传变异和遗传结构
八个银杏群体的遗传变异系数Fst为0.093,表明群体间的遗传变异很小,这可能是由于群体间的平均基因流较高(Nm=2.427,表3)。AMOVA的结果揭示了区域间(西南和东部)、区域内和群体内群体间的显著遗传差异(表5),但变异集中在群体内。总的来说,3%的变异发生在地理区域之间,10%和87%的变异分别发生在区域内和群体内的群体之间。
表3、22个微卫星位点的多态性信息。
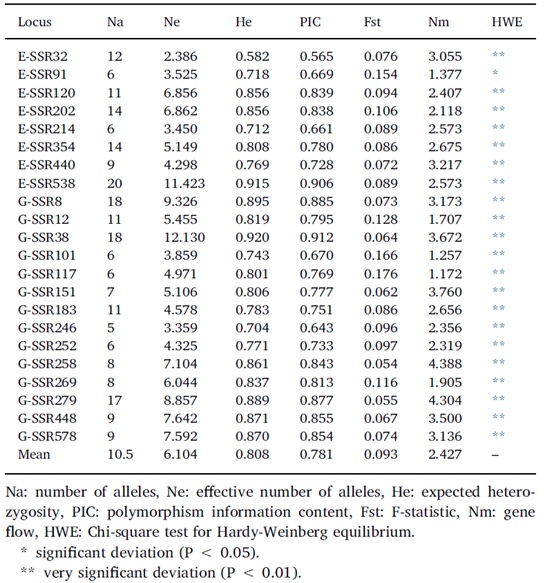
表5、来自8个种群的216个个体的AMOVA分析。
为了分析银杏群体的遗传结构,基于贝叶斯聚类模型分析了群体间的亲缘关系。根据DK的结果,当DK处于最大值时,最佳的K值是2(图3)。
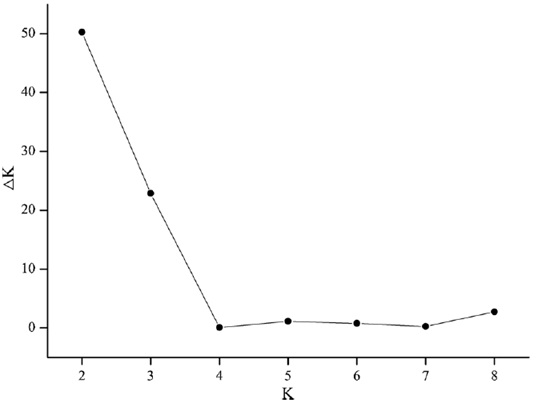
图3、聚类数(K)与相应的DK之间的关系图。
当K为2时,来自六个古代群体和两个栽培群体的216个样本的遗传信息来自两个差异祖先群体(图4a)。在K= 2时,中国西南部五个古代群体(PX、DY、MJ、FG和WC)的大部分遗传信息来自同一个祖先群体(绿色),而中国东部的TM和两个栽培群体(NJ和PZ)的遗传信息主要来自另一个祖先群体(红色)。在K = 3和4时,来自东方的三个群体的大多数遗传信息仍然来自相同的祖先群体,这表明
两个栽培种群(NJ和PZ)的起源与种群TM密切相关。然而,在西南部的五个古代群体中发现了基因差异。在五个古老的种群中,WC和PX种群具有丰富的遗传变异和起源的遗传信息。DY和MJ群体中的大多数遗传信息来源相似,与WC和PX群体也密切相关。另外,群体FG中的遗传信息是WC和PX群体中遗传信息的混合,表明群体FG的形成受两个古老群体的影响。
结果表明,8个种群的216个样本可分为2个类群(I组和II组),这与种群结构分析的模式一致。东部的TM、NJ和PZ三个种群的大部分遗传信息属于第一组,而西北部的五个古种群的大部分遗传信息属于第二组。此外,PCoA的结果进一步验证了系统发育树和种群结构分析结果(图4-C)。总之,根据遗传数据,这六个古代和两个栽培种群的起源明显不同,这些种群被分为西南组(PX、WC、DY、MJ和FG)和东部组(TM、NJ和PZ)。
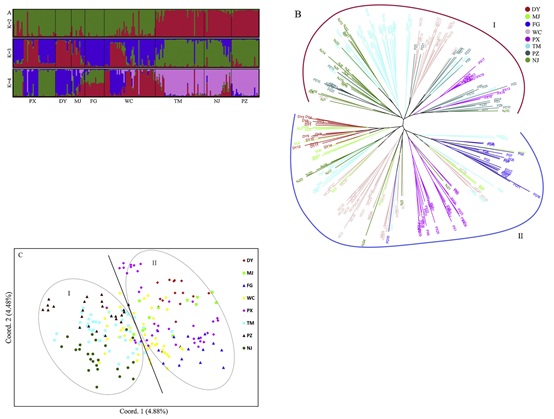
图4、A)基于贝叶斯聚类模型的216个样本群体遗传结构,K = 2-4。B)基于Nei’s遗传距离的邻居连接树。C) PCoA图。结果表明,样品可分为两类。
稀有等位基因和特异等位基因
在种群扩张过程中,低频等位基因比高频等位基因更容易丢失,这使得低频等位基因成为种群起源研究的重要基础。一般来说,原始群体比正常群体含有更多稀有等位基因和(或)特异等位基因。在216个样本中检测到的231个等位基因的频率范围从0.0023到0.6319,平均为0.0952。22个SSR基因座的最小等位基因频率在0.0023-0.0370之间,平均为0.0144。为了研究群体间的遗传变异,分析了在两个群体中检测到的稀有等位基因和仅在一个群体中检测到的特异等位基因(图5)。在22个SSR位点检测到18个稀有等位基因和17个特异等位基因。8个SSR位点具有稀有等位基因,其中279、183和354比其他位点含有更多的稀有等位基因。10个SSR位点中分布有特异的等位基因,在32个位点和8个位点分别检测到4个和3个等位基因。除DY群体外,其他群体都含有稀有等位基因,WC (8)和TM (9)群体比其他群体(3–4)含有更多稀有等位基因。四个群体(WC、TM、PZ和NJ)具有独特的等位基因,而群体TM具有最独特的等位基因(8),其次是新泽西州(6)、华盛顿(2)和PZ (1)。值得注意的是,尽管西南方的WC种群几乎没有特异的等位基因,但西南方的五个种群有14个等位基因在东部种群中未检测到(表6),这表明西南方的古种群包含特异的遗传信息。
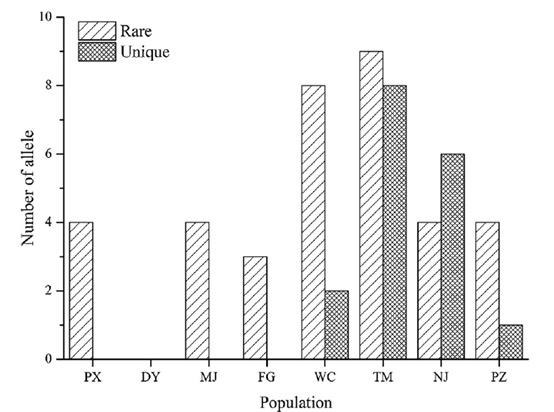
图5、8个银杏群体中稀有和独特等位基因的数量。
表6、西南5个群体中14个等位基因的频率。
4、讨论及结论
遗传多样性作为衡量一个物种适应环境变化能力的重要指标,受到许多因素的影响,如选择、遗传漂变、迁移和繁殖系统。研究银杏种群的遗传多样性,为银杏资源的保护和利用提供理论参考。
中国西南部位于青藏高原东南部和云贵高原,高原形成天然屏障,有效的减缓了冰期西伯利亚寒流对该地区的影响。由于其稳定的环境和多样的地貌,该地区拥有中国最丰富的生物多样性。本研究样本来自中国两个公认的潜在避难区,具有遗传多样性,等位基因罕见且独特。因此,这些树木是重要的遗传资源,可用于次级代谢产物的树木育种过程。然而,除FG和TM种群外,各种群的雌雄树比例均呈现不同程度的不平衡,雌树数量是雄树数量的2-6倍。DY、FG和MJ种群中的古树不到20棵,这些种群中的古树数量远远低于其他古种群。此外,22个SSR位点的所有等位基因均明显偏离哈迪-温伯格平衡,所有古代群体均表现出纯合子过量。因此,西南和东部的古银杏种群受到人类活动和自然因素的影响,应更加重视这两个地方的古银杏保护。
这项研究的结果证实,分布在两个潜在避难所(西南和东部)的古银杏种群具有高度的遗传多样性。两地样品间也发现了遗传变异,西南地区的种群起源明显不同,而东部种群起源明显。在冰河时期,东部的种群总数和西南部的种群总数是两个独立的避难所,中国可能还有其他避难所。此外,古代种群受到人类活动和自然因素的影响,应该进一步保护。该研究不仅为银杏避难所的研究提供了分子证据,也为古树的开发利用提供了理论指导。
关于天昊:
天昊生物可以提供SSR分子标记的一代毛细管电泳检测,并且基于二代高通量测序技术开发出SSRseqTM专利技术, 可以根据客户项目需求,提供不同数量样本和位点的高性价比SSR检测服务。

我们期待成为您SSR分型检测的优质服务合作伙伴,欢迎联系我们具体咨询!邮箱:techsupport@geneskies.com 电话:400-065-6886