2020年1月份10x Genomics平台单细胞转录组文章共计33篇(数据来源于10x官网),我们精选其中10篇文章总结了研究内容供大家阅读,文末汇总了其它文章的相关信息。
《一》

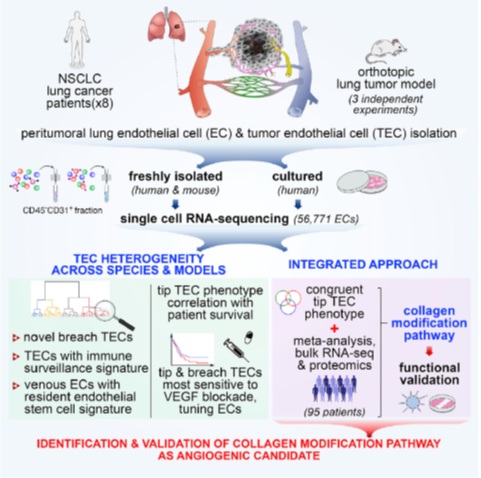
整合基因表达图谱分析方法发现了肺肿瘤内皮细胞异质性及血管生成的候选基因
摘要:患者、物种(人/小鼠)和模型(体内/体外)之间肺肿瘤内皮细胞(TEC)表型的异质性在单细胞水平上的研究仍然很少。我们用单细胞RNA测序了来自人(1例大细胞癌、4例鳞癌、3例腺癌,未经治疗)/小鼠肺肿瘤、正常肺及培养的人肺TECs的56,771个内皮细胞(MACS CD45-CD31+),并检测到17种已知的和16种以前未被识别的表型,包括推断的调节免疫监视的TECs。我们将典型的尖端(tip)TECs分解为已知的迁移尖端和假定的基底膜重塑破坏(breach)表型。Tip TEC特征与患者生存期相关,而Tip /breach TECs对血管内皮生长因子阻断最敏感。只有tip TECs在物种/模型间是一致的,并共享保守的标记。综合分析不同人肿瘤的单细胞测序数据、正交多组学和meta分析数据,通过功能分析验证,确定胶原修饰是一种候选的血管生成途径。
《二》

染色质比对和单细胞免疫分析确定慢性淋巴细胞白血病中依鲁替尼反应的时间动态
摘要:布鲁顿酪氨酸激酶(BTK)抑制剂依鲁替尼为慢性淋巴细胞白血病(CLL)患者提供了有效的治疗,尽管该疾病存在广泛的异质性。为了明确潜在的调控动态,我们分析了依鲁替尼治疗CLL患者的高分辨率时间进程,结合免疫表型、单细胞转录组分析和染色质比对。我们确定了一个一致的调控程序:在CLL细胞中开始时NF-κB结合大幅减少,紧随其后的是谱系定义(lineage-defining)的转录因子活性降低, CLL细胞的身份侵蚀,以及静止样基因特征出现。我们观察了病人对病人执行该程序速度的变化,并基于治疗前病人样本预测了病人对依鲁替尼反应的特定动态。总之,我们的研究描述了治疗性抑制CLL中B细胞受体信号的时间依赖性的细胞、分子和调控效应,并为基于表观基因组/转录组的治疗监测建立了一个广泛适用的方法。
《三》

人和小鼠单细胞核转录组测序显示阿尔茨海默症中依赖和不依赖TREM2的细胞反应
摘要:胶质细胞与阿尔茨海默病(AD)的发病机制有关。小胶质细胞表面的髓细胞触发受体2(TREM2)的变异增加了AD风险,而疾病相关的小胶质细胞(DAM)的激活依赖于AD小鼠模型中的TREM2。我们通过单细胞核RNA测序研究了5XFAD小鼠和人类AD中与AD病理和TREM2相关的基因表达变化。我们证实了基于Trem2的DAM的存在,并在小鼠中发现了一个以前未发现的Serpina3n+C4b+反应性少突胶质细胞群。有趣的是,在人类AD中明显存在显著不同的神经胶质表型。小胶质细胞的特征提示IRF8驱动的反应性小胶质细胞在周围神经损伤中的作用。少突胶质细胞信号表明轴突髓鞘化和对神经变性的代谢适应能力受损。星形胶质细胞的特征表明与神经元的代谢协调减弱。值得注意的是,小胶质细胞的反应表型在TREM2-R47H和TREM2-R62H突变携带者中比在非携带中更不明显,表明小鼠和人类AD中均需要TREM2,尽管存在明显的物种特异性差异。
《四》

整合人眼表达数量性状位点(eQTL)和单细胞图谱信息发现年龄相关性黄斑变性致病基因
摘要:年龄相关性黄斑变性(AMD)是视力下降的主要原因。为了更好地了解该病的发病机制,并在GWAS位点中识别AMD风险的致病基因,我们呈现了一个完整的人类视网膜和视网膜色素上皮(RPE)数据库。我们的数据库包括来自129个供体的黄斑和非黄斑RNA测序,一个包括黄斑特异性视网膜和RPE/脉络膜的全基因组表达数量性状位点(eQTL)数据集,以及来自人类视网膜和RPE的单核RNA-seq(NucSeq)数据,超过10万个细胞的亚型分辨率。利用NucSeq,我们发现RPE细胞中丰富的表达AMD候选基因。根据AMD风险和眼睛eQTL的遗传相关性信号的共定位,确定了15个可能的AMD致病基因,包括TSPAN10和TRPM1基因。这些结果证明了我们的人眼数据库在阐明眼部疾病的遗传途径和潜在治疗靶点方面的价值。
《五》

重建的单细胞命运轨迹定义人PSC诱导的末梢肺祖细胞分化过程中的谱系可塑性窗口
摘要:肺泡上皮2型细胞(AEC2s)是兼性祖细胞,负责维持肺泡的一生,但很难从患者中分离出来。我们在体外从人类多能干细胞(PSCs)中诱导AEC2s,并使用带有慢病毒条形码的时间进展的单细胞RNA测序来分析其与胎儿和成年AEC2基准相比的分化动力学。我们观察到细胞命运分岔轨迹作为原始肺祖细胞在体外分化,一些后代达到他们的AEC2命运目标,而其他的分化到选择性的非肺内胚层命运。我们建立了一个连续状态隐马尔可夫模型来识别信号的时间和类型,如过度兴奋的Wnt反应,这些信号会导致一些早期的多能NKX2-1+祖细胞失去肺命运。最后,我们发现这种最初的发育可塑性是可调节的,并随着时间的推移而减弱,最终导致PSC诱导的AEC2s表现出稳定的表型和几乎无限的自我更新能力。
《六》

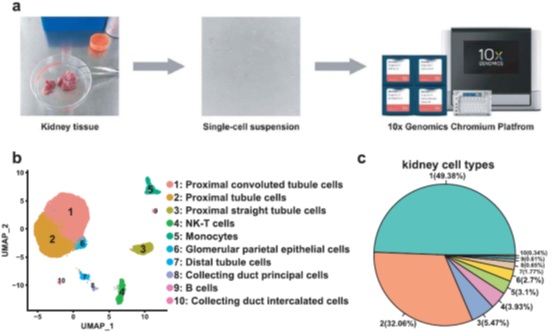
人肾脏单细胞测序
摘要:对正常人体肾脏进行全面的细胞解剖,对于解决肾脏疾病和肾癌的细胞起源问题至关重要。某些肾脏疾病可能是细胞类型特异性的,尤其是肾小管细胞。为了研究人类肾脏的分类和转录组信息,我们快速获得了肾脏的单细胞悬浮液,并进行了单细胞RNA测序(scRNA-seq)。这里,我们展示了来自三个人类肾脏捐赠者的23366个高质量细胞的scRNA-seq数据。在这个数据集中,我们显示了10类正常的人类肾细胞。由于单细胞转录组信息的高质量,近端小管(PT)细胞被分为三种亚型,收集导管细胞被分为两种亚型。总的来说,我们的数据为肾细胞生物学和肾脏疾病的研究提供了可靠的参考。
《七》

整个哺乳动物心脏的单细胞核测序
摘要:细胞水平的分析对于扩展我们对复杂组织如哺乳动物心脏的理解是必不可少的。单核测序(snRNA-seq)允许探索细胞组成和细胞特征,不存在单细胞测序的主要障碍。我们使用snRNA-seq首次对整个成年哺乳动物心脏进行了研究。单核定量和聚类可准确呈现细胞类型,显示出24个不同的聚类,其中内皮细胞(28.8%)、成纤维细胞(25.3%)和心肌细胞(22.8%)构成了主要的细胞群。一个额外的RNA速度分析使我们能够研究转录动力学,并被用来可视化细胞类型成熟和新生细胞状态之间的转变。我们用不同的标记物鉴定了心肌细胞亚群。例如,Hand2os1的表达将未成熟的心肌细胞与分化的心肌细胞群体区分开来。此外,我们发现了一个细胞群,包括内皮细胞标志物以及与心肌细胞功能明确相关的标志物。我们的速度数据支持了这一观点,即该群体正处于从内皮细胞样表型向心肌细胞样表型的转化过程中。综上所述,我们首次报道了对整个成年哺乳动物心脏的测序,提供了真实的细胞类型分布和RNA速度动力学的相互关系。
《八》

人类青春期期间睾丸发育的动态转录组细胞图谱
摘要:人类睾丸在青春期经历了剧烈的发育和结构变化,包括体细胞的增殖和成熟,以及精子形成的开始。为了表征这一未曾研究的过程,我们分析了4名青春期期间男孩的约10000个睾丸细胞的单细胞转录组数据,并将其与婴儿和成人的数据进行比较。在青春期,未分化的精原细胞在配子形成之前依次扩展和分化。值得注意的是,我们鉴定了睾丸间质和髓细胞的一种常见的青春期前的祖细胞,并描述了控制青春期分化的候选因素。此外,青春期前的塞尔托利氏细胞表现出两种不同的转录状态,它们在青春期向一个可替代的单一成熟群体趋同之前的代谢特征也不同。通过对睾酮抑制的变性女性睾丸的单细胞分析进一步强调了睾酮在塞尔托利氏细胞成熟、抗菌肽分泌和精原细胞分化中的作用。综上所述,我们的人类睾丸发育转录图谱提供了多种关于发育变化和伴随男性青春期的关键因素的见解。
《九》

肿瘤进化进程中基因组事件驱动着胰腺癌转录表型
摘要:胰脏腺癌是一种侵袭性极强的疾病。这种疾病的异质性的基础已经被证明是难以解决的,因为差的肿瘤细胞结构和广泛的基因组不稳定性。为了解决这个问题,从纯化的原发和转移性肿瘤的上皮细胞中产生了一个完整的基因组和转录组数据集。转录组分析表明,分子亚型是由肿瘤内混合亚群驱动的基因表达连续体的产物,这已被单细胞分析证实。整合的全基因组分析发现,分子亚型与KRAS突变和GATA6等基因的特定拷贝数变异有关。通过绘制肿瘤遗传历史,四倍体化成为这些事件背后的一个关键突变过程。综上所述,这些数据支持了这样一个前提,即肿瘤中基因组变异的集合导致了分子亚型,而疾病异质性是由于在进展过程中持续的基因组不稳定性造成的。
《十》

发现人类β细胞中LIF响应、可复制的亚群
摘要:在胚胎发生过程中形成的β细胞团在胎儿和产后早期发育过程中被细胞复制放大。此后,β细胞功能成熟,通过低复制率维持其细胞团。对于那些在成年期复制的少数β细胞,不知道复制是如何开始的,也不知道这是否发生在β细胞的一个特定亚群中。我们利用YAP过表达系统诱导干细胞来源的β细胞的复制,并通过单细胞RNA测序发现了白血病抑制因子(LIF)通路的上调。LIF通路的激活可诱导人β细胞在体内和体外的复制。LIF受体的表达被限制在一个转录独特的人类β细胞的亚群中,该亚群的增殖能力增强。这项研究描绘了新的基因网络,控制着LIF响应的,复制能力强的人类β细胞的复制。